生物化學與分子生物學/DNA復制的延長階段以及參與的酶和蛋白質分子
| 醫學電子書 >> 《生物化學與分子生物學》 >> DNA的生物合成 >> DNA的復制 >> DNA復制的延長階段以及參與的酶和蛋白質分子 |
| 生物化學與分子生物學 |
|
|
|
DNA 的復制實際上就是以 DNA 為模板在 DNA聚合酶 作用下,將游離的四種脫氧 單核苷酸(dATP,dGTP,dCTP,dTTP, 簡寫為 dNTP) 聚合成 DNA 的過程。
這是一個非常復雜的 酶促反應,需要許多種酶和 蛋白質 參與,現分別敘述它們在 DNA 復制中作用。
(一)DNA 的聚合反應和 DNA 聚合酶
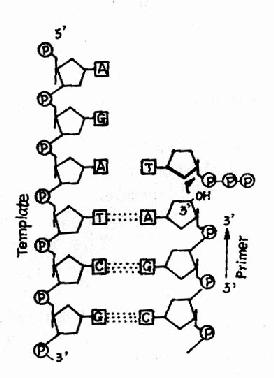
圖 16-9 DNA 聚合酶的作用
1957 年,Arthur kornberg 首次在 大腸桿菌 中發現 DNA 聚合酶Ⅰ,(DNa polymerase Ⅰ, 簡寫 DNA polⅠ) 后來又相繼發現了 DNA 聚合酶Ⅱ和 DNA 聚合酶Ⅲ。(DNa polymerase Ⅱ,Ⅲ,簡寫 DNA polⅡ,DNA polⅢ) 實驗證明大腸桿菌中 DNA 復制的主要過程靠 DNa polⅢ起作用,而 DNA polⅠ和 DNA polⅡ在 DNA 錯配的校正和修復中起作用。見表 16-1。
這種酶的共同性質是:①需要 DNA 模板,因此這類酶又稱為依賴 DNA 的 DNA 聚合酶 (DNa dependent DNA polymerase, DDDP)。②需要 RNA 或 DNA 做為 引物(primer),即 DNA 聚合酶不能從頭 催化DNA 的起始。③催化 dNTP 加到引物的 3′桹 H 末端,因而 DNA 合成的方向是 5′→3′。圖 16-9。④三種 DNA 聚合酶都屬于 多功能酶,它們在 DNA 復制和修復過程的不同階段發揮作用。由于 DNA 聚合酶Ⅰ是研究得最清楚而且代表了其他 DNA 聚合酶的基本特點,所以我們著重介紹 DNa polⅠ的作用并指出另外二種 DNA pol 的特殊性:
1.DNA 聚合酶Ⅰ:
DNA polⅠ是由一條 多肽 鏈組成,分子量 為 109KD。酶 分子 中含有一個 Zn++,是聚合活性必須的。
大腸桿菌每個 細胞 中約有 400 個酶分子,每個酶分子每分鐘在 37℃下能催化 667 個 核苷酸 參入到 DNA 鏈中,用 枯草桿菌蛋白酶 可將此酶水介成兩個片段,大片段分子量為 76KD,通常稱為 klenow 片段,小片段為 34KD。大小片段具有不同的 酶活性。
(1)DNA 聚合酶的 5′→3′聚合活性:
這是 DNA 聚合酶最主要的活性,按模板 DNA 上的核苷酸順序,將互補的 dNTP 逐個加到引物 RNA3′桹 H 末端,并促進 3′桹 H 與 dNTP 的 5′桺 O4 形成 磷酸二酯鍵,酶的 專一性 表現為新進入的 dNTP 必須與模板 DNA堿基配對 時才有催化作用,5′→3′聚合活性存在于 klenow 片段上 (圖 16-9 和圖 16-10)。
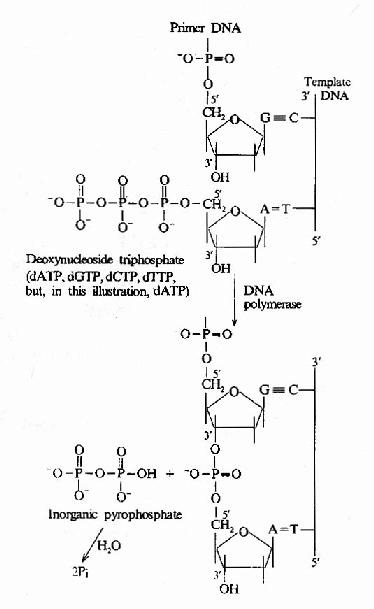
圖 16-10 DNA 聚合酶催化的 DNA 鏈延長
(2)DNA 聚合酶的 3′→5′外切 核酸酶 活性:
這種酶活性的主要功能是從 3′→5′方向識別并切除 DNA 生長鏈末端與模板 DNA 不配對而游離的核苷酸,這種功能稱為校對功能,這是保證其聚合作用的正確性不可缺少的,因此對于 DNA 復制中極高的保真性是至關重要的。
(3)DNA 聚合酶的 5′→3′外切核酸酶活性:
這種酶活性是從 DNA 鏈的 5′端向 3′末端水解已配對的核苷酸,本質是切斷磷酸二酯鍵,每次能切除 10 個核苷酸。因此,這種酶活性在 DNA 損傷的修復中可能起重要作用,對完成的 DNA 片段去除 5′端的 RNA 引物也是必須的。
DNA polⅠ的 5′→3′聚合活性和 5′→3′外切酶活性協同作用,可以使 DNA 一條鏈上的切口從 5′→3′方向移動,這種反應叫做缺刻平移 (nick translation),利用此反應可在體外對 DNA 片段進行 放射性 磷 (α-32PdNTP) 的標記制成 探針(probe),進行 核酸 的 分子雜交 實驗,是現代分子 生物學 的一項重要技術 (圖 16-11)。
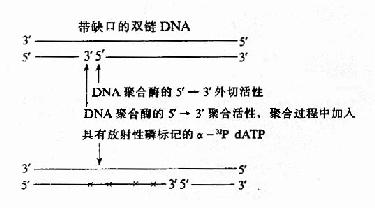
圖 16-11 缺刻平移標記 DNA 探針
許多實驗證實 DNA polⅠ并不是 DNA 復制過程中的主要酶,它的作用主要與 DNA 損傷后的修復有關。
2.DNA 聚合酶Ⅱ(DNA polⅡ)
此酶分子量為 120KD,每個細胞約有 100 個酶分子,但活性只有 DNa polⅠ的 5%,它具有 5′→3′聚合活性和 3′→5′外切活性,而沒有 5′→3′外切活性,它的作用可能與 DNA 損傷修復有關。
3.DNA 聚合酶Ⅲ(DNA polⅢ)
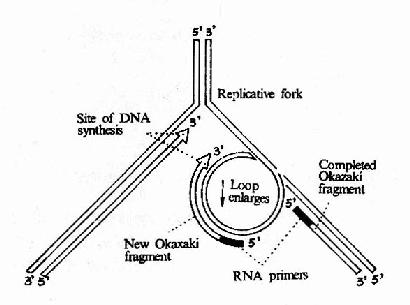
圖 16-12 DNA 聚合酶Ⅲ催化先導鏈和隨從的合成
這是在 DNA 復制過程中起主要作用的聚合酶,它是由一個 亞基 組成的蛋白質分子,其分子量>600kDa 整個酶分子形成一個不對稱的 二聚體,每個大腸桿菌細胞中只有 10?0 個酶分子,但催化 dNTP 參入 DNA 鏈的速率卻是最快的,約為 9000 核苷酸 / 每分鐘 / 每個酶分子。這也證明 DNa polⅢ是 DNA 復制過程中主要發揮作用的酶。在大腸桿菌 染色體DNA 進行復制時,DNA 聚合酶Ⅲ全酶 并不是單獨起作用的,而是與 引發體,介 鏈酶 等構成一個復制體 (replisome)。由于復制體的存在,先導鏈和隨從鏈可以同時復制。DNa polⅢ是由多亞基組成的不對稱二聚體,它可能同時負責先導鏈和隨從鏈的復制,在φ×174 的復制中觀察到引發體總是伴隨著 DNA 嚕噗 (loop) 的存在。圖 16?2 可以看到,由于隨從鏈的模板 DNA 在 DNA 聚合酶Ⅲ全酶上繞轉了 180°而形成一個嚕噗,因此崗崎片段的合成方向能夠與先導鏈的合成方向以及復制體移動方向保持一致。
隨著 DNA polⅢ向前移動,先導鏈的合成逐漸延長的同時,崗崎片段也在不斷延長,這一嚕噗也在不斷擴大。當崗崎片段合成到前一個片段的 5′端時,這一大嚕噗就釋放出來,由于 復制叉 向前移動又可將另一部分隨從鏈的模板置換出來,由引發體合成新的引物,然后再形成一個小的嚕噗,進行新的崗崎片段的合成。由此模型不難看出:隨從鏈的合成需要 周期性 的引發,因此其合成進度總是與 前導鏈 相差一個崗崎片段的長度。崗崎片段完成后,其 5′端的 RNA 引物由 DNa polⅠ的 5′→3′外切酶活性切除,由此造成的空隙再由 DNA polⅠ的 5′→3′聚合活性催化 dNTP 得到填補。所以 DNA 的復制是在 DNa polⅢ和 DNApolⅠ互相配合下完成的。
下面列表說明三種大腸桿菌 DNA 聚合酶的特性
表 16-1 大腸桿菌 DNA 聚合酶特征
| DNA 聚合酶Ⅰ | DNA 聚合酶Ⅱ | DNA 聚合酶Ⅲ | |
| 分子量 | 109KD | 120KD | >600KD |
| 每個細胞中的分子數 | 400 | 17-100 | 10-20 |
| 5′→3′聚合活性 | + | + | + |
| 37℃轉化率核苷酸數/酶分子. 分鐘 | 600 | 30 | 30,000 |
| 5′→3′外切活性 | + | - | - |
| 3′→5′外切活性 | + | + | + |
| 切刻平移活性 | + | - | - |
| 對 dNTP 親和力 | 低 | 低 | 高 |
| 功能 | 修復 | 不詳 | 復制 |
| 去除引物 | |||
| 填補空缺 |
真核生物DNA 聚合酶
真核生物 DNA 聚合酶有α、β、γ、δ及ε。它們的基本特性相似于大腸桿菌 DNA 聚合酶,其主要活性是催化 dNTP 的 5′→3′聚合活性,基本特征見表 16-2。
表 16-2 真核生物 DNA 聚合酶
| α | β | γ | δ | ε | |
| 亞基數 | 4 | 4 | 4 | 2 | 5 |
| 分子量(KD) | >250 | 36-38 | 160-300 | 170 | 256 |
| 細胞內定位 | 核 | 核 | 線粒體 | 核 | 核 |
| 5′→3′聚合活性 | + | + | + | + | + |
| 3′→5′外切活性 | - | - | - | - | - |
| 功能 | 復制、引發 | 修復 | 復制 | 復制 | 復制 |
真核細胞 在 DNA 復制中起主要作用的是 DNA polα,主要負責染色體 DNA 的復制。DNa polβ的模板特異性是具有缺口的 DNA 分子,被認為它與 DNA 修復 有關。DNa polγ在線粒體 DNA 的復制中起作用。DNA polδ不但有 5′→3′聚合活性,而且還具有 3′→5′外切酶活性,據認為真核生物 DNA 復制是在 DNa polα和 DNA polδ協同作用下進行的,前導鏈的合成靠 DNA polδ催化,并且還需要一種 細胞周期 調節因子椩鮒誠赴絲乖?proliferatingcell nucleus antigen, PCNA) 參與。而隨從鏈的合成靠 DNA polα和 引發酶 配合作用完成。
(二) 與 超螺旋 松馳有關的酶:
DNA 復制從起始點開始向一個方向復制時,局部的 DNA 雙鏈必須打開,主要靠 解鏈酶 的作用,打開后的 單鏈 還需要單鏈 結合蛋白 與其結合,在復制叉向前移動時造成其前方 DNA 分子所產生的正超螺旋,必須由 拓撲異構酶 來解決。下面分別介紹它們的作用。
拓撲異構酶 (topoisomerase) 是一類改變 DNA 拓撲性質的酶。在體外可催化 DNA 的各種拓撲異構化反應,而在生物體內它們可能參與了 DNA 的復制與 轉錄。在 DNA 復制時,復制叉行進的前方 DNA 分子部分產生有正超螺旋,拓撲酶可松馳超螺旋,有利于復制叉的前進及 DNA 的合成。DNA 復制完成后,拓撲酶又可將 DNA 分子引入超螺旋,使 DNA 纏繞、折疊,壓縮以形成 染色質。DNA 拓撲異構酶有Ⅰ型和Ⅱ型,它們廣泛存在于 原核生物 及真核生物中。表 16-3
表 16-3 大腸桿菌和真核 生物 中的拓撲異構酶
| 類型 | 作用 | 對超螺旋的作用 |
| Ⅰ型拓撲異構酶 | ||
| 大腸桿菌 | 切開一股 DNA 鏈 | 松馳負超螺旋 |
| 真核生物 | 切開一股 DNA 鏈 | 松馳正,負超螺旋 |
| Ⅱ型拓撲異構酶 | ||
| 大腸桿菌 | 切開二股 DNA 鏈 | 構馳正超螺旋; |
| 依賴 ATP | 引入負超螺旋, | |
| 解環連等 | ||
| 真核生物 | 切開二股 DNA 鏈 | 松馳正超旋, |
| 依賴 ATP | 但不能引入負超螺旋 |
拓撲異構酶Ⅰ(TopoⅠ) 的主要作用是將環狀雙鏈 DNA 的一條鏈切開一個口,切口處鏈的末端繞螺旋軸按照松馳超螺旋的方向轉動,然后再將切口封起來。這就使 DNA 復制叉移動時所引起的前方 DNA 正超螺旋得到緩解,利于 DNA 復制叉繼續向前打開。拓撲異構酶Ⅰ除上述作用外,對環狀 單鏈 DNA 還有打結或解結作用,對環狀雙鏈 DNA 的環連或解連以及使環狀單鏈 DNA 形成環狀雙鏈 DNA 都有作用 (圖 16-13)。
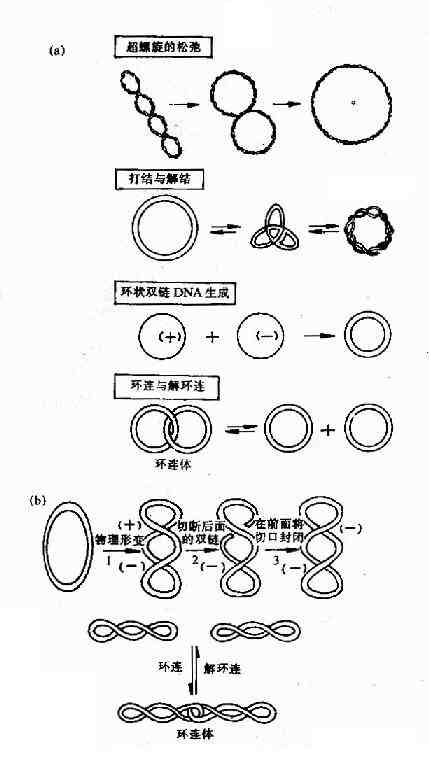
圖 16-13 拓撲酶Ⅰ及Ⅱ的作用特點
(a) 大腸桿菌拓撲酶Ⅰ催化的 4 種拓撲異構化作用 (b) 拓撲酶Ⅱ的作用
拓撲異構酶Ⅱ(TopoⅡ) 是在大腸桿菌中發現的,曾被稱為 旋轉酶(gyrase),它們作用特點是切開環狀雙鏈 DNA 的兩條鏈,分子中的部分經切口穿過而旋轉,然后封閉切口,TopoⅡ還可使 DNA 分子從超螺旋狀態轉變為松馳狀態,此反應不需要 ATP 參與。DNA 復制完成后,TopoⅡ在 ATP 參與下,DNA 分子從松馳狀態轉變為負超螺旋。此外,TopoⅡ催化的拓撲異構化反應還有環連或解環連,以及打結或解結。
| 關于“生物化學與分子生物學/DNA復制的延長階段以及參與的酶和蛋白質分子”的留言: | |
|
目前暫無留言 | |
| 添加留言 | |