生物化學與分子生物學/真核基因表達調控的特點
| 醫學電子書 >> 《生物化學與分子生物學》 >> 基因表達調控 >> 真核基因表達調控 >> 真核基因表達調控的特點 |
| 生物化學與分子生物學 |
|
|
|
盡管我們現在對真核基因表達調控知道還不多,但與原核生物比較它具有一些明顯的特點。
(一)真核基因表達調控的環節更多
如前所述,基因表達是基因經過轉錄、翻譯、產生有生物活性的蛋白質的整個過程。同原核生物一樣,轉錄依然是真核生物基因表達調控的主要環節。但真核基因轉錄發生在細胞核(線粒體基因的轉錄在線粒體內),翻譯則多在胞漿,兩個過程是分開的,因此其調控增加了更多的環節和復雜性,轉錄后的調控占有了更多的分量。圖19-13扼要地列出真核基因表達的各個可能的環節。
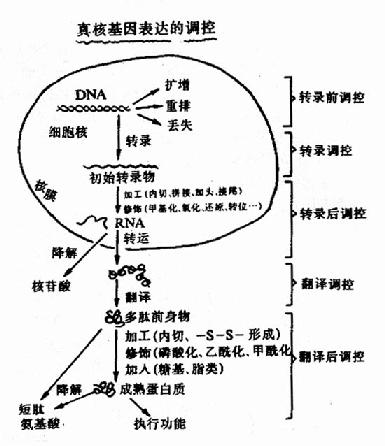
圖19-13 真核生物基因表達調控的可能環節
圖19-13總結了以前章節敘述過的基因表達過程,并作了一些新補充。圖中標出了真核細胞在分化過程中會發生基因重排(gene rearrangement),即胚原性基因組中某些基因會再組合變化形成第二級基因。例如編碼完整抗體蛋白的基因是在淋巴細胞分化發育過程中,由原來分開的幾百個不同的可變區基因經選擇、組合、變化,與恒定區基因一起構成穩定的、為特定的完整抗體蛋白編碼的可表達的基因。這種基因重排使細胞可能利用幾百個抗體基因的片段,組合變化而產生能編碼達108種不同抗體的基因,其中就有復雜的基因表達調控機理。
此外,真核細胞中還會發生基因擴增(geneamplification),即基因組中的特定段落在某些情況下會復制產生許多拷貝。最早發現的是蛙的成熟卵細胞在受精后的發育過程中其rRNA基因(可稱為rDNA)可擴增2000倍,以后發現其他動物的卵細胞也有同樣的情況,這很顯然適合了受精后迅速發育分裂要合成大量蛋白質,需要有大量核糖體。又如MTX(methotrexate)是葉酸的結構類似物,一些哺乳類細胞會對含有利用葉酸所必需的二氫葉酸還原酶(dihydrofolate reductase, DHFR)基因的DNA區段擴增40?00倍,使DHFR的表達量顯著增加,從而提高對MTX的抗性。基因的擴增無疑能夠大幅度提高基因表達產物的量,但這種調控機理至今還不清楚。
(二)真核基因的轉錄與染色質的結構變化相關
真核基因組DNA絕大部分都在細胞核內與組蛋白等結合成染色質,染色質的結構、染色質中NA和組蛋白的結構狀態都影響轉錄,至少有以下現象:
1.染色質結構影響基因轉錄 細胞分裂時染色體的大部分到間期時松開分散在核內,稱為常染色質(euchromatin),松散的染色質中的基因可以轉錄。染色體中的某些區段到分裂期后不像其他部分解旋松開,仍保持緊湊折疊的結構,在間期核中可以看到其濃集的斑塊,稱為異染色質(heterochromatin),其中從未見有基因轉錄表達;原本在常染色質中表達的基因如移到異染色質內也會停止表達;哺乳類雌體細胞2條X染色體,到間期一條變成異染色質者,這條X染色體上的基因就全部失活。可見緊密的染色質結構阻止基因表達。
2.組蛋白的作用 早期體外實驗觀察到組蛋白與DNA結合阻止DNA上基因的轉錄,去除組蛋基因又能夠轉錄。組蛋白是堿性蛋白質,帶正電荷,可與DNA鏈上帶負電荷的磷酸基相結合,從而遮蔽了DNA分子,妨礙了轉錄,可能扮演了非特異性阻遏蛋白的作用;染色質中的非組蛋白成分具有組織細胞特異性,可能消除組蛋白的阻遏,起到特異性的去阻遏促轉錄作用。
發現核小體后,進一步觀察核小體結構與基因轉錄的關系,發現活躍轉錄的染色質區段,有富含賴氨酸的組蛋白(H1組蛋白)水平降低,H2A.H2B組蛋白二聚體不穩定性增加、組蛋白乙酰化(acetylation)和泛素化(ubiquitination),以及H3組蛋白巰基化等現象,這些都是核小體不穩定或解體的因素或指征。轉錄活躍的區域也常缺乏核小體的結構。這些都表明核小體結構影響基因轉錄。
3.轉錄活躍區域對核酸酶作用敏感度增加 染色質DNA受DNase Ⅰ作用通常會被降解成00、400……bp的片段,反映了完整的核小體規則的重復結構。但活躍進行轉錄的染色質區域受DNase Ⅰ消化常出現100-200bp的DNA片段,且長短不均一,說明其DNA受組蛋白掩蓋的結構有變化,出現了對DNase Ⅰ高敏感點(hypersensitive site)。這種高敏感點常出現在轉錄基因的5′側區(5′flanking region)、3′末端或在基因上,多在調控蛋白結合位點的附近,分析該區域核小體的結構發生變化,可能有利于調控蛋白結合而促進轉錄。
4.DNA拓撲結構變化 天然雙鏈DNA的構象大多是負性超螺旋。當基因活躍轉錄時,RNA聚合酶轉錄方向前方DNA的構象是正性超螺旋,其后面的DNA為負性超螺旋。正性超螺旋會拆散核小體,有利于RNA聚合酶向前移動轉錄;而負性超螺旋則有利于核小體的再形成。
5.DNA堿基修飾變化 真核DNA中的胞嘧啶約有5%被甲基化為5甲基胞嘧啶(5methylcytidine,m5C),而活躍轉錄的DNA段落中胞嘧啶甲基化程度常較低。這種甲基化最常發生在某些基因5′側區的CpG序列中,實驗表明這段序列甲基化可使其后的基因不能轉錄,甲基化可能阻礙轉錄因子與DNA特定部位的結合從而影響轉錄。如果用基因打靶的方法除去主要的DNA甲基化酶,小鼠的胚胎就不能正常發育而死亡,可見DNA的甲基化對基因表達調控是重要的。
由此可見,染色質中的基因轉錄前先要有一個被激活的過程,但目前對激活機制還缺乏認識。
(三)真核基因表達以正性調控為主
真核RNA聚合酶對啟動子的親和力很低,基本上不依靠自身來起始轉錄,需要依賴多種激活蛋白的協同作用。真核基因調控中雖然也發現有負性調控組件,但其存在并不普遍;真核基因轉錄表達的調控蛋白也有起阻遏和激活作用或兼有兩種作用者,但總的是以激活蛋白的作用為主。即多數真核基因在沒有調控蛋白作用時是不轉錄的,需要表達時就要有激活的蛋白質來促進轉錄。換言之:真核基因表達以正性調控為主導。
| 關于“生物化學與分子生物學/真核基因表達調控的特點”的留言: | |
|
目前暫無留言 | |
| 添加留言 | |